INTRODUCTION
Hearing is one of the major senses and like vision is important for distant warning and communication. It can be used to alert, to communicate pleasure and fear. It is a conscious appreciation of vibration perceived as sound. In order to do this, the appropriate signal must reach the higher parts of the brain. The function of the ear is to convert physical vibration into an encoded nervous impulse. It can be thought of as a biological microphone. Like a microphone the ear is stimulated by vibration: in the microphone the vibration is transduced into an electrical signal, in the ear into a nervous impulse which in turn is then processed by the central auditory pathways of the brain. The mechanism to achieve this is complex. This chapter will deal mainly with the ear, first its structure and then its function, for it is the ear that is mainly at risk from hazardous sounds.
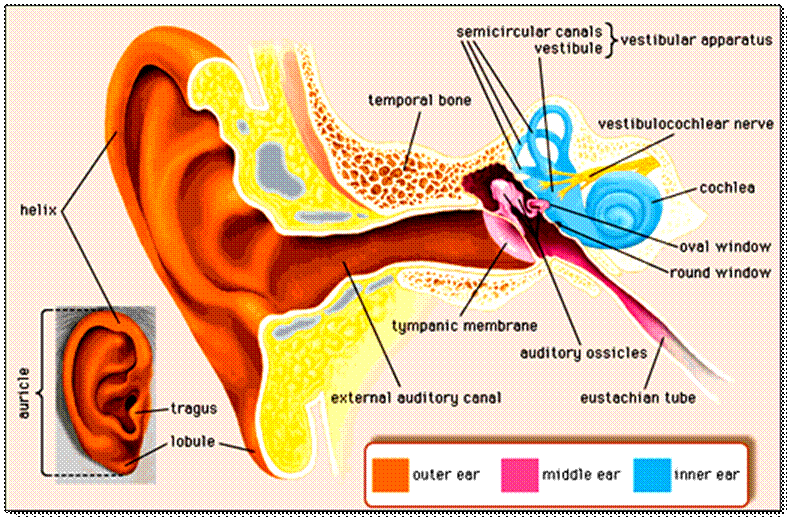
The ear is made up of three parts: the outer, middle, and inner ear. All three parts of the ear are important for detecting sound by working together to move sound from the outer part through the middle and into the inner part of the ear. Ears also help to maintain balance.
OUTER EAR
The outer ear transmits sound to the tympanic membrane. The pinna, that part which protrudes from the side of the skull, made of cartilage covered by skin, collects sound and channels it into the ear canal. The pinna is angled so that it catches sounds that come from in front more than those from behind and so is already helpful in localizing sound. Because of the relative size of the head and the wavelength of audible sound, this effect only applies at higher frequencies. In the middle frequencies the head itself casts a sound shadow and in the lower frequencies phase of arrival of a sound between the ears helps localize a sound. The ear canal is about 4 centimetres long and consists of an outer and inner part. The outer portion is lined with hairy skin containing sweat glands and oily sebaceous glands which together form ear wax. Hairs grow in the outer part of the ear canal and they and the wax serve as a protective barrier and a disinfectant. Very quickly however, the skin of the ear canal becomes thin and simple and is attached firmly to the bone of the deeper ear canal, a hard cavity which absorbs little sound but directs it to the drum head (eardrum or tympanic membrane) at its base. The outer layer of the drumhead itself is formed of skin in continuity with that of the ear canal.
In life, skin sheds and is continuously renewing. Ear canal skin grows like a fingernail fromthe depths to the exterior so that the skin is shed into the waxy secretions in the outer part and falls out. This is the reason for not using cotton buds to clean the ear canal because very frequently they merely push the shed skin and wax deep into the canal, impacting it and obstructing hearing. The ear canal has a slight bend where the outer cartilaginous part joins the bony thin skinned inner portion, so that the outer part runs somewhat backwards and the inner part somewhat forwards. This bend is yet another part of the protective mechanism of the ear, stopping foreign objects from reaching the tympanic membrane. However it means that to inspect the tympanic membrane from the outside, one must pull the ear upwards and backwards. The tympanic membrane separates the ear canal from the middle ear and is the first part of the sound transducing mechanism. Shaped somewhat like a loudspeaker cone (which is an ideal shape for transmitting sound between solids and air), it is a simple membrane covered by a very thin layer of skin on the outside, a thin lining membrane of the respiratory epithelium tract on the inner surface and with a stiffening fibrous middle layer. The whole membrane is less than a 1/10th of millimetre thick. It covers a round opening about 1 centimetre in diameter into the middle ear cavity. Although the tympanic membrane is often called the ear drum, technically the whole middle ear space is the ear drum and the tympanic membrane the drum skin.
MIDDLE EAR
The middle ear is an air filled space connected to the back of the nose by a long, thin tube called the Eustachian tube. The middle ear space houses three little bones, the hammer, anvil and stirrup (malleus, incus and stapes) which conduct sound from the tympanic membrane to the inner ear. The outer wall of the middle ear is the tympanic membrane, the inner wall is the cochlea. The upper limit of the middle ear forms the bone beneath the middle lobe of the brain and the floor of the middle ear covers the beginning of the great vein that drains blood from the head, the jugular bulb. At the front end of the middle ear lies the opening of the Eustachian tube and at its posterior end is a passageway to a group of air cells within the temporal bone known as the mastoid air cells. One can think of the middle ear space shaped rather like a frying pan on its side with a handle pointing downwards and forwards (the Eustachian tube) but with a hole in the back wall leading to a piece of spongy bone with many air cells, the mastoid air cells. The middle ear is an extension of the respiratory air spaces of the nose and the sinuses and is lined with respiratory membrane, thick near the Eustachian tube and thin as it passes into the mastoid. It has the ability to secret mucus. The Eustachian tube is bony as it leaves the ear but as it nears the back end of the nose, in the nasopharynx, consists of cartilage and muscle. Contracture of muscle actively opens the tube and allows the air pressure in the middle ear and the nose to equalize.
Sound is conducted from the tympanic membrane to the inner ear by three bones, the malleus, incus and stapes. The malleus is shaped like a club; its handle is embedded in the tympanic membrane, running from its centre upwards. The head of the club lies in a cavity of the middle ear above the tympanic membrane (the attic) where it is suspended by a ligament from the bone that forms the covering of the brain. Here the head articulates with the incus which is cone shaped, with the base of the cone articulating with the head of the malleus, also in the attic. The incus runs backwards from the malleus and has sticking down from it a very little thin projection known as its long process which hangs freely in the middle ear. It has a right angle bend at its tip which is attached to the stapes(stirrup), the third bone shaped with an arch and a foot plate. The foot plate covers the oval window, an opening into the vestibule of the inner ear or cochlea, with which it articulates by the stapedio-vestibular joint.
INNER EAR
The inner ear is made up of two labyrinths – the bony labyrinth and the membranous labyrinth, both of which are encased by a protective bony shell called the otic capsule. The bony labyrinth can be divided into 3 distinct areas: the vestibule; the three semi-circular canals and the cochlea. Within these lie the separate areas of the membranous labyrinth: the saccule; the utricle; the semicircular ducts and the cochlear duct or scala media. Functionally, the inner ear can be divided into two systems. The vestibular system which is concerned with the mechanisms of balance comprises the semicircular canals, the vestibule and the membranous sacs and ducts they enclose. The snail-like cochlea forms part of the auditory system, with the oval window of the bony labyrinth situated at its base to allowing transmission of sound from the outer and middle ears.
The cochlea
The cochlear duct is a spiral-shaped bony tube surrounding a central core called the modiolus. In humans the spiral is two and three quarter turns in length, in the mouse it is one and three quarters. The cochlear duct is divided along its length into three chambers, the scala vestibuli, the scala tympani and the scala media. The scala vestibuli and scala tympani form the outer layers of the duct and both contain perilymph which is high in sodium ion (Na+ ) concentration and low in potassium ion (K+ ) concentration like most other extracellular fluids. Between them lies the scala media, a cavity which is triangular in cross section and filled with endolymph high in K+ concentration and low in Na+ concentration, similar to the intracellular fluids of neurons.
The division between the scala media and the scala tympani below it is formed by the basilar membrane. This structure is composed of connective tissue, with a distinct basement membrane separating it from the overlying organ of Corti. Above the scala media the boundary is formed by Reissner’s membrane, while the external wall attaches to the wall of the bony labyrinth and is termed the lateral wall. This wall includes the spiral ligament and stria vascularis which play important roles in recycling endolymph and maintaining the ion gradients within the scala media.
The organ of Corti
The organ of Corti is the highly specialised sensory patch of the cochlea and is composed of sensory hair cells and supporting cells. It rests on the basilar membrane and extends through the length of the cochlea within the scala media (Figure 1.4). Hair cells can be distinguished by the bundles of stereocilia on their apical surfaces and form two distinct types; the inner hair cells (IHCs) and outer hair cells (OHCs). The IHCs are arranged in a single row towards the modiolar edge of the organ of Corti, while the OHCs are ordered in three parallel rows (sometimes up to four rows in the apex) slightly further away from the central core of the cochlea. IHCs have a flask-shaped cellular body and a centrally located nucleus and account for 90- 95% of the afferent innervation leading to the cochlear nucleus (Spoendlin 1972). The OHCs are longer and cylindrical in shape, and have an abundant efferent nerve supply. However, the most noticeable difference between the two types of hair cells is the arrangement of the stereocilia on their surface. The IHCs have their stereocilia arranged in a slightly crescent manner, whilst the stereocilia on OHCs are arranged in a distinctive “v” shape in the apex, widening to form a “w” shape in the base. In general the stereocilia of IHCs are longer and of larger diameter than those of the OHCs, which tend to be short and narrow. The stereocilia of both hair cell types form a “staircase” gradient with the tallest stereocilia at the outer edge of the cell. Stereocilia are composed of tightly packed actin filaments, making them relatively strong and rigid. Between the rows of stereocilia exist tip links and numerous lateral links which preserve the integrity of the bundle and also mean that the stereocilia in a bundle all move together. The deflection of the stereocilia bundle towards the tallest stereocilia row produces an excitatory response, as discussed later.
The sensory hair cells in the organ of Corti are surrounded by a variety of different supporting cells. Lying between the single row of IHCs and the first row of OHCs are two rows of microtubule-packed inner and outer pillar cells. As well as separating the two hair cell types, these cells give the organ of Corti rigidity along its length. The upper ends of the pillar cells form a plate called the reticular lamina which forms the division between the endolymph of the scala media and the perilymph of the scala tympani below. Surrounding the pillar cells are phalangeal cells. Inner phalangeal cells surround the inner IHCs, while outer phalangeal cells (also called Deiter’s cells) support the OHCs, forming a cup at their base and extending fine microtubule-filled processes towards the reticular lamina which serve to separate the three rows of OHCs. Hensen’s cells form the outer border external to the OHC rows, while inner border cells are situated between the row of IHCs and the modiolus.
The tallest stereocilia of the OHCs are embedded in the fibrous and gelatinous tectorial membrane. This structure is anchored to the spiral limbus and forms a flap which covers the organ of Corti. The IHCs are not embedded in this matrix, but instead fit loosely into a groove on its under surface (Pickles 1988).
Auditory transduction
Auditory transduction refers to the transformation of the tiny vibrational movements of the basilar membrane and the organ of Corti which rests upon it into electrical impulses. The sound waves that enter the cochlea via the outer and middle ear at the oval window cause the displacement of endolymph in the scala media which is accommodated by the round window. This movement of the fluid surrounding the basilar membrane causes it to be displaced, with the frequency of the sound determining the pattern of movement. Sounds of a low frequency result in a vibration pattern which peaks in the apex of the cochlea, while high frequency tones produce more vibration in the basal region. This arrangement is described as tonotopic organisation. The movement of the basilar membrane in turn causes displacement of the stereocilia bundles on the surfaces of the sensory hair cells. This displacement occurs because the tectorial membrane in which their tips are embedded has a different pivot point from the basilar membrane which forms their base. Thus when the membranes are displaced, the stereocilia also undergo displacement.
Mechanical stimuli applied to a hair bundle elicit electrical responses by causing transduction channels to open or close. When the stereocilia bundle is deflected towards the tallest row of stereocilia, tension in the tip-links which connect adjacent stereocilia exerts force on the transducer channels and causes more of them to open, thus allowing an influx of extracellular K+ and Ca+ which causes depolarisation of the cell. A stimulus which brings about a movement of the stereocilia towards the short edge of the bundle has the opposite effect, causing the transducer channels to close and hyperpolarizing the cell. The degree of polarisation in turn affects the rate of release of a synaptic transmitter which causes the transmission of a pattern of action potentials to the brain. These are carried along an afferent nerve fibre which contacts the base of the cell, and encodes information regarding the intensity, time course and frequency of the sound stimulation.
The vestibular system
The vestibular system is concerned with the perception of static and dynamic equilibrium which is necessary for balance and spatial orientation. Static equilibrium relates to the position of the body in relation to gravity, while dynamic equilibrium is concerned with the maintenance of body position in response to movement. It comprises the semicircular canals and the otolithic organs, the saccule and the utricle, as well as the endolymphatic sac and duct (Figure 1.3). The endlymphatic duct joins the endolymphatic sac to the utricle and plays a role in fluid homeostasis; the anatomy and function of the remaining components are described below.
The semicircular canals
Within the bony semicircular canals lie the lateral, anterior and posterior semicircular ducts (Figure 1.3). Both ends of each canal join to the vestibule, while the rostral end opens out into an ampulla, an expanded cavity containing a sensory patch known as the crista ampullaris. Each of these is an elongated epithelial structure situated on a ridge of supporting tissue arising from the wall of the ampulla, with its surface populated by three distinct cell types. The first of these are supporting cells, characterised by a tall columnar shape and short microvilli on the apical surface. The other two cell types are sensory hair cells, which are classified as type I or type II depending on their morphology and innervation. Type I cells are flask shaped, with an afferent nerve chalice and some efferent nerve fibres surrounding the baso-lateral region. Type II cells are cylindrical in shape, and possess both efferent and afferent innervation at their base. Both type I and type II cells possess stereocilia on their apical surface. These are stiff actin-filled microvilli which are organised into a “staircase” shape, with taller projections at the back, gradually decreasing in height towards the front. The stereocilia of the hair cells project into a cone-shaped gelatinous membrane called the cupula. Each cell also has a single kinocilium – a cilium containing the classic 9+2 arrangement of microtubules at its core – which is always located on the utricular side of the hair cell.
The semicircular ducts and their associated cristae are responsible for the detection of changes in the direction and rate of movement of the head, known as rotational acceleration and deceleration. In order to account for all planes of movement, the superior and posterior canals are in vertical planes perpendicular to each other, while the lateral canal is in a near-horizontal plane. Each crista is located perpendicular to the plane of the duct with which it is associated. Rotation of the head in the plane of that semicircular canal causes movement of the endolymph within it, which in turn displaces the cupula. This causes movement of the embedded stereocilia bundles towards or away from the kinocilium, resulting in depolarising or hyperpolarising receptor potentials respectively and the release of a neurotransmitter at varying rates, initialising neural impulses towards the central vestibular system.
The otolithic organs
The utricle and saccule are dilated regions of the central vestibule, each containing a sensory patch termed a macula. The two maculae are located perpendicular to each other and are composed of supporting cells and type I and type II sensory hair cells similar to those found in the cristae. The stereocilia of the hair cells are embedded in a gelatinous layer of glycoprotein called the otolithic membrane, a sheet-like structure which is thought to be secreted by the supporting cells. The upper surface of this membrane contains minute crystalline bodies composed of calcium carbonate and protein which are known as otoconia. The maculae are responsible for the maintenance of static equilibrium – the detection of gravity – which is of particular importance when other cues as to spatial orientation are lacking, such as when the eyes are closed, or in the dark or under water. Movement of the head causes the otolithic membrane, weighed down by the otoliths, to move in relation to the hair cells beneath them, giving rise to displacement of the stereocilia. As in the cristae and the cochlea, this causes increased excitation when the movement is towards the kinocilium and reduced excitation when the movement is away from the kinocilium.